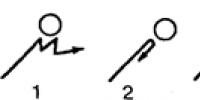Cilia and flagella: a concise description, structure and role in cells. Flagella structure and diversity Habitats of the simplest unicellular organisms
A more complex form is the movement carried out with the help of flagella and cilia. The flagellar form of movement is characteristic of the class of flagellates.
Flagella are the thinnest outgrowths of the body. The number of them in different species is different - from one to many tens and even hundreds.
Each flagellum originates from a small basal granule called the blepharoplast, located in the cytoplasm. Thus, the part of the flagellum directly adjacent to the basal grain passes inside the cytoplasm (it is called the root thread), and then passes through the pellicle to the outside. The mechanism of flagellar movement is different in different species. In most cases, it comes down to rotational movement. The flagellum describes the figure of a cone, with its apex facing the place of its attachment. The greatest mechanical effect is achieved when the angle formed by the top of the cone is 40-46°. The speed of movement is different, it varies in different species between 10 and 40 revolutions per second. The simplest is, as it were, "screwed" into the liquid medium surrounding it.
Often, the rotational movement of the flagellum is combined with its undulating movement. Usually, during translational motion, the body of the protozoan itself rotates around a longitudinal axis.
The outlined scheme is valid for most single-flagellated forms.
In polyflagellates, the movement of the flagella may be of a different nature, in particular, the flagella may be in the same plane without forming a cone of rotation.
Rice. Cross section of the flagella of Pseudotrichonympha (drawing taken from an electron microscope photograph). Nine peripheral double fibrils and a central pair of fibrils are visible in each flagellum.
Electron microscopic studies of recent years have shown that the internal ultramicroscopic structure of the flagella is very complex. Outside, the flagellum is surrounded by a thin membrane, which is a direct continuation of the most superficial layer of ectoplasm - the pellicle. The internal cavity of the flagellum is filled with cytoplasmic contents. Eleven finest filaments (fibrils) run along the longitudinal axis of the flagellum, which are often double (Fig.). These fibrils are always arranged regularly. Nine of them (simple or double) lie along the periphery, forming together, as it were, a cylinder. Two fibrils occupy a central position. To get an idea of the size of all these formations, it is enough to say that the diameter of the peripheral fibrils is about 350 A (angstrom). An angstrom is a unit of length equal to 0.0001 microns, and a micron is equal to 0.001 mm. These structures, insignificant in size, have become available for study due to the introduction of the electron microscope into microscopic technology.
The functional significance of flagellum fibrils cannot be considered definitively elucidated. Apparently, some of them (probably peripheral) play an active role in the motor function of the flagellum and contain special protein molecules that can contract, while others are supporting elastic structures that have a supporting value.
Cilia serve as organelles for the movement of ciliates. Usually their number in each individual is very large and is measured in several hundred, thousands and even tens of thousands. The mechanism of movement of cilia is somewhat different from that of flagella.
Each eyelash makes rowing movements. She quickly and forcefully bends to one side, and then slowly straightens.
The joint action of a large number of cilia, the beating of which is coordinated, causes a rapid forward movement of the protozoan.
Each ciliary cilium, as shown by the latest research, is a complex formation, in its structure corresponding to the flagellum. At the base of each cilium, there is always the so-called basal grain (otherwise, kinetosoma), an important part of the ciliary apparatus.
In many ciliates, individual cilia are connected to each other, forming structures of a more complex structure (membranella, cirri, etc.) and more efficient mechanical action.
Some highly organized protozoa (ciliates, radiolarians) are characterized by another form of movement - contraction. The body of such protozoa is able to quickly change its shape, and then return to its original state again.
The ability to quickly contract is due to the presence in the body of the simplest special fibers - myonemes - formations similar to the muscles of multicellular animals.
Some protozoans also have other forms of movement.
The mobility of many protists is provided by the presence of flagella or cilia. Both of them are set up in the same way. Flagella in both protozoa and flagella or ciliary cells of multicellular animals and plants are always only a part of the locomotor system of the cell, which consists of a kinetosome (or centriole), a flagellum (or undulipodium) and root outgrowths of the kinetosome (or its derivatives (derivatives)). In addition to movement in the water column, flagella and cilia are used for temporary or permanent attachment to the substrate or for creating food flows of water when feeding on suspended particles.
Flagellum is a tubular outgrowth of the cell surface, surrounded by a membrane that serves as a continuation of the membrane that covers the entire cell. It contains a bundle of protein fibrils, the so-called axoneme. An axoneme or axial filament is a microtubule formation that consists of two central microtubules surrounded by a ring of nine pairs (doublets) of microtubules, consisting of subfibrils tightly fused to each other. The fine structure of the flagella of all eukaryotic organisms is surprisingly uniform in its main features.
The most important element of the flagellar system is the basal body or kinetosome. This is a cylinder, the walls of which are formed by nine groups of microtubules, united in threes (triplets). Most often, two kinetosomes are present in the cell, located approximately at right angles to each other. One or two flagella depart from them. The kinetosome does not float in the cytoplasm by itself, since it is fixed by a system of roots.
Modern ideas about the Protista system are largely based on the structure of the flagellum and its derivatives. The wide distribution of flagella and cilia among them makes it possible to compare almost all taxa with each other, and also makes it possible to use additional characters of the flagellar apparatus, the number of which is already approaching 100, in taxonomy and phylogeny. Many structural features of flagellates, including the shape of the body, are determined by the presence of this peculiar system in them.
The number of flagella, their relative and absolute length, the place and method of attachment of the flagella, the nature of their movement, their orientation are very diverse in different groups, but are constant within individual groups of related organisms.
Usually, 4 morphotypes of flagellates are distinguished.
Isoconts have from 2 to 8 flagella of equal length, directed in one direction, with the same ways of beating. These include most of the motile cells of green algae.
Anisoconts have 2 flagella of unequal length, directed in one direction, differing in the way they beat. Such flagella are characteristic of colorless flagellates.
At heterocont there are 2 flagella of unequal length (one directed forward, the other backward), differing in the way they beat. They are also characteristic of motile cells of algae, and of the so-called zoospore fungi, and of colorless flagellates.
Stephanokonty have a corolla of flagella at the anterior end of the cell. This is characteristic of polyflagellated gametes and zoospores of some green algae.
Single-flagellated forms are usually not distinguished into a special group. Many of them are considered as individuals that have lost their flagellum for the second time, since the vast majority have one more flagellaless kinetosome.
The main function of the flagellum is movement. In the active work of the flagellum, peripheral microtubules and their handles, which have ATPase activity, are the driving principle. Central microtubules have a reference value. The forms of movement of the flagellum are different, but usually it is a helical movement, allowing the flagellate to "screw" into the water, making up to 40 revolutions per second. In ciliates and polyflagellated protists, the movement of cilia is organized according to the type of metachronal waves. Flagella and cilia are often used for nutrition as well. Among the flagellates, there are species that spend most of their life cycle in an attached state. During this period, the flagellum loses its usual function of movement and turns into an attachment organelle, a stalk or a leg. Another function of the flagellum is that, with its movements, it cleans the surface of the body from small foreign particles adhering to it.
Endoplasmic organelles
In the endoplasm of protists there are one or more nuclei, as well as all organelles and structures characteristic of a eukaryotic cell: ER, ribosomes, Golgi apparatus, mitochondria, peroxisomes, hydrogenosomes, plastids (in autotrophic protists), lysosomes, digestive vacuoles. Some protists also have organelles specific only to them.
Extrusoms. These organelles are special vacuoles surrounded by a membrane, which in mature extruses usually contacts the plasmalemma. In response to various external stimuli (mechanical, chemical, electrical, etc.), they throw out their contents. In their structure, these are mucopolysaccharides (complex compounds of carbohydrates with proteins). There are 10 different types of extrusoms known. Some contain poisonous substances that can immobilize and kill the victim (protozoa and other small organisms). Others perform a protective function or facilitate movement by secreting mucus.
Plastids. Plastids are present in phototrophic and related protists and are represented by chloroplasts and leucoplasts. The main pigments of chloroplasts are chlorophylls. Different groups of phototrophic protists are characterized by certain sets of chlorophylls. Of the secondary pigments in algae, carotenes and xanthophylls are found, which in high concentrations can mask green chlorophyll and give chloroplasts a variety of colors from yellow-green to reddish-brown.
The Golgi apparatus has been found in almost all studied species of protists. Most often, the Golgi apparatus is located next to the nucleus and is represented by one or more stacks of flat cisterns (dictyosomes) surrounded by small vesicles. However, the Golgi apparatus in some protists is formed by single cisterns. The absence of dictyosomes is usually interpreted as a primitive feature. However, the absence of dictyosomes in modern protists cannot unambiguously indicate their primitiveness, since the formation and disassembly of dictyosomes largely depend on external influences on the cell (for example, a decrease in the oxygen concentration in the environment) or on physiological rearrangements of the protist itself (transition to encystation).
Lysosomes and other organelles and inclusions. In protist cells, as in the cells of multicellular animals, there are lysosomes. These cytoplasmic bodies in the form of small vesicles (primary lysosomes) are formed in the Golgi apparatus. Digestive hydrolytic enzymes are localized in them. Secondary lysosomes, or digestive vacuoles, are well expressed only in heterotrophic protists feeding by phagocytosis.
The endoplasm of various protists contains more or less reserve nutrients used in metabolic processes. Most often, these are various polysaccharides (glycogen, starch, amyloplectin, etc.), often lipids and other fatty inclusions. The amount of reserve substances depends on the physiological state of the protozoan, the nature and amount of food, on the stage of the life cycle, and varies widely. However, some large groups of protists store specific substances. For example, Euglenoids store paramyl, which is not found in other protists.
Body protozoan is made up of cytoplasm and one or more cores. The nucleus is surrounded by a double membrane and contains chromatin, which includes deoxyribonucleic acid (DNA), which determines the genetic information of the cell. Most protozoa have a vesicular nucleus with a small amount of chromatin collected along the periphery of the nucleus or in an intranuclear body, the karyosome. Micronuclei of ciliates are massive nuclei with a large amount of chromatin. Common cell components of most protozoa include mitochondria and the Golgi apparatus.
Surface bodies of amoeboid forms(sarcodal, as well as some stages of the life cycle of other groups) is covered with a cell membrane about 100 A thick. Most protozoa have a denser, but elastic shell, the pellicle. The body of many flagellates is covered with a periplast formed by a series of longitudinal fibrils fused with the pellicle. Many protozoa have special supporting fibrils, such as the supporting fibril of the undulating membrane in trypanosomes and Trichomonas.
Thick and hard shells have resting forms of protozoa, cysts. Shell amoeba, foraminifera and some other protozoa are enclosed in houses or shells.
Unlike cells of a multicellular organism the cell of the simplest is a complete organism. To perform the diverse functions of the body in the body of the simplest, structural formations, organelles, can specialize. According to their purpose, the organelles of protozoa are divided into organelles of movement, nutrition, excretion, etc.
Very diverse protozoan movement organelles. Amoeboid forms move through the formation of protrusions of the cytoplasm, pseudopodia. This type of movement is called amoeboid and is found in many groups of protozoa (sarcode, asexual forms of sporozoans, etc.). Flagella and cilia serve as special organelles for movement. Flagella are characteristic of the class of flagellates, as well as gametes of representatives of other classes. They are few in most forms (from 1 to 8). The number of cilia, which are the organelles of the movement of ciliates, can reach several thousand in one individual. An electron microscope study showed that flagella and cilia in Protozoa, Metazoa and plant cells are built according to a single type. Their basis is a bundle of fibrils, consisting of two central and nine paired, peripheral ones.
tourniquet surrounded by a shell which is an extension of the cell membrane. Central fibrils are present only in the free part of the tourniquet, and peripheral fibrils go deep into the cytoplasm, forming a basal grain - blepharoplast. The tourniquet can be connected to the cytoplasm for a considerable distance by a thin membrane - an undulating membrane. The ciliary apparatus of ciliates can reach considerable complexity and differentiate into zones that perform independent functions. The cilia often coalesce in groups, forming spikes and membranellae. Each cilium starts from a basal grain, a kinetosome, located in the surface layer of the cytoplasm. The collection of kinetosomes forms the infracilia. Knetosomes reproduce only by dividing in two and cannot arise anew. With a partial or complete reduction of the flagellar apparatus, the infracilia remains and subsequently gives rise to new cilia.
The movements of protozoa are very diverse, and in this type of protozoa there are methods of locomotion that are completely absent in multicellular animals. This is a peculiar way of movement of amoebas with the help of "transfusion" of plasma from one part of the body to another. Other representatives of the protozoa, gregarines, move in a kind of “reactive” way - by secreting mucus from the rear end of the body, “pushing” the animal forward. There are also protozoa that passively hover in the water.
However, most protozoa move actively with the help of special structures that produce rhythmic movements - flagella or cilia. These effectors are plasma outgrowths that perform oscillatory, rotational, or wave-like movements. Flagella, long hair-like outgrowths, are possessed by the already mentioned primitive protozoa, which received their name due to this formation. With the help of flagella, the body of an animal (for example, euglena) is brought into a spiral translational motion. Some marine flagellates, according to the Norwegian scientist I. Trondsen, rotate when moving around an axis at a speed of up to 10 revolutions per second, and the translational speed can reach 370 microns per second. Other marine flagellates (from among dinoflagellates) develop speeds from 14 to 120 microns per second or more. A more complex effector apparatus is the cilia that cover the body of the ciliate in large numbers. As a rule, the ciliary cover is located unevenly, the cilia reach different lengths on different parts of the body, form ring-shaped seals (membranella), etc.
An example of such a complex differentiation is ciliates from the genus Stilonychia. The peculiar organelles of these animals allow them not only to swim, but also to "run" on a solid substrate, both forward and backward. It has been established that the coordination of these methods and directions of locomotion, as well as their "switching", is carried out by special mechanisms localized in three centers and two axes of excitation gradients in the cytoplasm.
Flagella and cilia are set in motion by contractions of myofibrils, which form fibers, myonemas, corresponding to the muscles of multicellular animals. In most protozoa, they are the main locomotor apparatus, and even the most primitive representatives of the flagellate type have them. Myonemes are arranged in a strict order, most often in the form of rings, longitudinal threads or ribbons, and in the higher representatives and in the form of specialized systems. Thus, the ciliate Caloscolex has special systems of myonemes of perioral membranella, pharynx, hindgut, a number of retractors of individual parts of the body, etc.
It is interesting to note that, as a rule, myonemas have a homogeneous structure, which corresponds to the smooth muscles of multicellular animals, but sometimes there are also transversely striated myonemas, comparable to the striated muscles of higher animals. All contractile fibrils serve to perform fast movements of individual effectors (in protozoa, needle-like outgrowths, tentacle-like formations, etc.). Complex systems of myonemes allow the protozoa to produce not only simple contractile movements of the body, but also quite a variety of specialized locomotor and non-locomotor movements.
In those protozoa that do not have myonemes (amoebas, rhizopods, sporozoans, with one exception, and some other protozoa), contractile movements occur directly in the cytoplasm. So, when the amoeba moves in the outer layer of the cytoplasm, in the ectoplasm, genuine contractile processes occur. It was even possible to establish that these phenomena take place every time in the "rear" (in relation to the direction of movement) part of the body of the amoeba.
Thus, even before the appearance of special effectors, the movement of the animal in space is accomplished by contractions. It is the contractile function, which is carried in protozoa by myonemes, and in multicellular muscles, that provided all the diversity and complexity of the motor activity of animals at all stages of phylogenesis.
Kinesis
Locomotion of the simplest is carried out in the form of kinesis - elementary instinctive movements. A typical example of kinesis is orthokinesis - translational movement with variable speed. If, for example, there is a temperature gradient (temperature difference) in a certain area, then the movement of the shoe will be the faster, the farther the animal is from the place with the optimum temperature. Consequently, here the intensity of the behavioral (locomotor) act is directly determined by the spatial structure of the external stimulus.
In contrast to orthokinesis, in klinokinesis there is a change in the direction of movement. This change is not purposeful, but is in the nature of trial and error, as a result of which the animal eventually enters the zone with the most favorable parameters of stimuli. The frequency and intensity of these changes (hence, the angle of rotation) depend on the intensity of the (negative) stimulus (or stimuli) acting on the animal. With the weakening of the strength of this stimulus, the intensity of klinokinesis also decreases. Thus, here, too, the animal reacts to the stimulus gradient, but not by increasing or decreasing the speed of movement, as in orthokinesis, but by turning the body axis, i.e. change in the vector of motor activity.
As we can see, the implementation of the most primitive instinctive movements - kinesis - is determined by the direct influence of intensity gradients of biologically significant external factors. The role of the internal processes occurring in the cytoplasm lies in the fact that they give the behavioral act the "first impetus", as in multicellular animals.
Orientation
We have already seen from examples of kinesis that gradients of external stimuli act in protozoa simultaneously as triggering and guiding stimuli. This is especially evident in clinokinesis. However, changes in the position of the animal in space are not yet truly orienting here, since they are non-directional. To achieve a full biological effect, clinokinetic, as well as orthokinetic, movements require additional correction, which allows the animal to more adequately navigate in its environment according to the sources of irritation, and not just change the nature of the movement under adverse conditions.
The orienting elements in representatives of the type under consideration and in other lower invertebrates at a given level of mental development are the simplest taxis. In orthokinesis, the orienting component - orthotaxis - manifests itself in a change in the speed of movement without changing its direction in the gradient of an external stimulus. In clinokineses, this component is called clinotaxis and manifests itself in a change in the direction of movement by a certain angle.
Rice. 26. Clinotaxis behavior of ciliates in a temperature gradient. Once in a warm zone (above), the infusoria goes to a cooler one (below) (according to Jennings)
As we already know, taxises are understood as genetically fixed mechanisms of spatial orientation of animal motor activity towards favorable (positive taxises) or away from unfavorable (negative taxises) environmental conditions. For example, negative thermotaxis are expressed in protozoa, as a rule, in the fact that they swim away from zones with a relatively high water temperature, less often from zones with a low temperature. As a result, the animal finds itself in a certain zone of thermal optimum (zone of preferred temperature). In the case of orthokinesis in a temperature gradient, negative orthothermotaxis provides a straight-line distance from unfavorable thermal conditions. If there is a clinokinetic reaction (Fig. 26), then clinotaxis provides a clear change in the direction of movement, thereby orienting random clinokinetic movements in the stimulus gradient (in our example, in the thermal gradient).
Often, clinotaxis manifests itself in rhythmic pendulum-like movements (in place or during movement) or in the spiral trajectory of a swimming animal. And here there is a regular rotation of the axis of the body of the animal (in multicellular animals it can be only a part of the body, for example, the head) at a certain angle.
Clinotaxis are also found when meeting with solid barriers. Here is an example of the corresponding behavior of ciliates (shoes). Having stumbled upon a solid barrier (or having fallen into a zone with other unfavorable environmental parameters), the shoe stops, the nature of the beating of the cilia changes, and the animal swims back a little. After that, the infusoria turns to a certain angle and swims forward again. This continues until she swims past the obstacle (or passes the unfavorable zone).

Rice. 27. Dependence of the behavior of ciliates on the angle of encounter with an obstacle (according to Rose). See text for explanations
According to new data (V. Rose), ciliates, however, do not always behave in this way. Clinotaxis are observed only when meeting an obstacle at an angle of 65–85° (Fig. 27, 1). If the meeting occurs at a right angle, the ciliate turns over through the transverse axis of the body and “bounces” back (Fig. 27, 2). If the animal encounters an obstacle along a tangent, it swims past, turning to the side (Fig. 27, 3).
Clinotaxis, although they are also found in more highly organized animals, belong to the primitive types of taxis. Protozoa do not have tropotaxis, since tropotaxis involves the presence of symmetrically located sensory organs.
In the examples given, the reactions of protozoa (in this case, ciliates) to temperature and tactile stimulus (touch) were described. It was, therefore, about thermo- and thigmotaxis, in the latter case - about negative thigmoclinotaxis, which occurs in response to strong tactile stimulation (contact with a solid surface of an object).
If, on the contrary, the shoe stumbles not on a hard obstacle, but on a soft object (for example, plant residues, filter paper), it reacts differently: with such weak tactile stimulation, the ciliate stops and is applied to this surface so that the maximum area of the body is in contact with object surface (positive thigmotaxis). A similar picture is also observed when other modalities affect the direction of movement, i.e. the positive or negative nature of the reaction depends on the intensity of the stimulus. As a rule, protozoa react positively to weak stimuli, negatively to strong ones, but in general, protozoa tend to avoid adverse effects rather than actively seek positive stimuli.
Returning to thigmotaxis, it is important to note that special receptors for tactile sensitivity were found in ciliates - tactile “hairs”, which are especially prominent at the anterior and posterior ends of the body. These formations do not serve to search for food, but only for tactile examination of the surfaces of objects that the animal encounters. Irritation of these organelles leads in the described example to the termination of the kinetic reaction.
A feature of the thigmotaxis reaction is that it often weakens and then stops after touching the object with the maximum surface of the body: the shoe adhering to the object begins to react to a different stimulus to an increasing extent and more and more separates from the object. Then, on the contrary, the role of the tactile stimulus increases again, and so on. As a result, the animal makes rhythmic oscillatory movements near the object.
The shoe also has a clearly expressed orientation in the vertical plane, which finds its expression in the tendency to swim upwards (negative geotaxis - orientation by the force of gravity). Since special organelles of gravitational sensitivity were not found in paramecium, it was suggested that the contents of the digestive vacuoles act in it like statocysts* of higher animals. The validity of this interpretation is confirmed by the fact that a shoe that swallowed metal powder in the experiment no longer floats up, but down, if a magnet is placed above it. In this case, the content of the vacuole (metal powder) no longer presses on its lower part, but, on the contrary, on its upper one, which, obviously, is the reason for the reorientation of the direction of animal movement by 180°
* Statocyst is an organ of sense of balance, the main part of which consists of a closed cavity with a statolith included in it - a “pebble” (mineral concretion).
In addition to the aforementioned taxis reactions, protozoa have also been established in response to chemical stimuli (chemotaxis), electric current (galvanotaxis), etc. Some of the protozoa react weakly to light, while in others this reaction is very clearly expressed. So, phototaxis appear in some species of amoebae and ciliates in a negative form, and the ciliate Blepharisma is even equipped with a photosensitive pigment, but, as a rule, ciliates do not respond to changes in lighting. It is interesting to note that in protozoans that react weakly to light, phenomena of summation of reactions to mechanical stimuli were noted if the latter were combined with light stimuli. This combination can increase the response eight times compared to the response to mechanical stimulation alone.
In contrast to ciliates, in many flagellates, especially in euglena, positive phototaxis is very clearly expressed. The biological significance of this taxis is beyond doubt, since the autotrophic nutrition of euglena requires solar energy. Euglena swims towards the light source in a spiral, at the same time, as already mentioned, rotating around its own axis. This is significant, since Euglena, like some other protozoans that react strongly and positively to light, have well-developed photoreceptors. These are pigment spots, sometimes even equipped with reflective formations that allow the animal to localize light rays. Moving towards the light source in the manner described, the euglena turns to it either the “blind” (dorsal) side or the “sighted” (abdominal) side. And every time the latter (with an unshielded section of the “eye”) turns out to be facing the light source, the trajectory of movement is corrected by turning at a certain angle towards this source. Consequently, the movement of the euglena towards the light is determined by positive photoclinotaxis, and in the case of exposure to two light sources, the alternating stimulation of the photoreceptor either on the left or on the right gives the movement of the euglena an outward resemblance to the tropotaxis behavior of bilaterally symmetrical animals with paired eyes (Fig. 28), which will be discussed below.
"Eyes" are also described in other flagellates. For example, a massive bright red eye has Collodictyon sparsevacuolata, a peculiar protist with 2–4 flagella and an amoeboid appearance, capable of moving quickly both with the help of flagella and with the help of prolegs. Photoreception is particularly complex in the flagellar Pauchetia (Dinoflagellata), which already has analogues of the essential parts of the eye of multicellular animals, the pigment spot is equipped not only with an opaque screen (analogue of the pigment membrane), but also with a translucent formation in the form of a spherical lens (analogue of the lens). Such an "eye" allows not only to localize light rays, but also to collect, to a certain extent focus them.

Rice. 28. The trajectory of the movement of euglena under the influence of two equally intense light sources (according to Buder)
Both prokaryotic and eukaryotic can contain structures known as cilia and flagella. These outgrowths on the surface of the cells help in their.
Features and functions
Cilia and flagella are outgrowths from certain cells necessary for cellular locomotion (movement). They also help move substances around the cells and guide them to the right places.
Cilia and flagella are formed from specialized groups of microtubules called basal bodies.
If the outgrowths are short and numerous, they are called cilia. If they are longer and less numerous (usually only one or two), they are called flagella.
Structure
Typically, cilia and flagella have a core consisting of microtubules connected to , arranged in a 9 + 2 pattern. The ring of nine microtubules has two special microtubules in its center that flex the cilia or flagella. This type of organization is found in the structure of most cilia and flagella.
Where do they meet?
Both cilia and flagella are found in many cell types. For example, the sperm of many animals, algae, and even ferns have flagella. Cilia can be found in cells in tissues such as the respiratory tract and the female reproductive tract.