Бактерии. Строение микроорганизмов Классификация вредных бактерий
Бактерии это самый древний организм на земле, а также самый простой в своем строении. Он состоит всего из одной клетки, которую можно увидеть и изучить только под микроскопом. Характерным признаком бактерий является отсутствие ядра, вот почему бактерии относят к прокариотам.
Некоторые виды образовывают небольшие группы клеток, такие скопления могут быть окружены капсулой (чехлом). Размер, форма и цвет бактерии сильно зависит от окружающей среды.
По форме бактерии различаются на: палочковидные (бациллы), сферические (кокки) и извитые (спириллы). Встречаются и видоизмененные – кубические, С-образные, звездчатые. Их размеры колеблются от 1 до 10мкм. Отдельные виды бактерий могут активно передвигаться при помощи жгутиков. Последние иногда превышают размер самой бактерии в два раза.
Виды форм бактерий
Для движения бактерии используют жгутики, количество которых бывает различное – один, пара, пучок жгутиков. Расположение жгутиков также бывает разным – с одной стороны клетки, по бокам или равномерно распределены по всей плоскости. Также одним из способов передвижения считается скольжение благодаря слизи, которой покрыт прокариот. У большинства внутри цитоплазмы есть вакуоли. Регулировка ёмкости газа в вакуолях помогает им двигаться в жидкости вверх или вниз, а также перемещаться по воздушных каналах почвы.
Ученые открыли более 10 тысяч разновидностей бактерий, но по предположениям научных исследователей в мире существует их более миллиона видов. Общая характеристика бактерий дает возможность определиться с их ролью в биосфере, а также изучить строение, виды и классификацию царства бактерий.
Места обитания
Простота строения и быстрота адаптации к окружающим условиям помогла бактериям распространиться в широком диапазоне нашей планеты. Они существуют везде: вода, почва, воздух, живые организмы – всё это максимально приемлемое место обитания для прокариотов.
Бактерии находили как на южном полюсе, так и в гейзерах. Они есть на океанском дне, а также в верхних слоях воздушной оболочки Земли. Бактерии живут везде, но их количество зависит от благоприятных условий. К примеру, большая численность видов бактерий проживает в открытых водоемах, а также почве.
Особенности строения
Клетка бактерии отличается не только тем, что в ней нет ядра, но и отсутствием митохондрий и пластид. ДНК данного прокариота находится в специальной ядерной зоне и имеет вид замкнутого в кольцо нуклеоида. У бактерии строение клетки состоит из клеточной стенки, капсулы, капсулоподобной оболочки, жгутиков, пили и цитоплазматичной мембраны. Внутреннее строение оформляют цитоплазма, гранулы, мезосомы, рибосомы, плазмиды, включения и нуклеоид.

Клеточная стенка бактерии выполняет функцию обороны и опоры. Вещества могут свободно протекать сквозь неё, благодаря проницаемости. Данная оболочка имеет в своем составе пектин и гемицеллюлозу. Некоторые бактерии выделяют особую слизь, которая может помочь защититься от пересыхания. Слизь формирует капсулу – полисахарид по химическому составу. В такой форме бактерия способна переносить даже очень большие температуры. Также она выполняет и другие функции, к примеру слипание с любыми поверхностями.
На поверхности клетки бактерии находятся тонкие белковые ворсинки – пили. Их может быть большая численность. Пили помогают клетке передавать генетический материал, а также обеспечивают слипание с другими клетками.
Под плоскостью стенки находится трехслойная цитоплазматичная мембрана. Она гарантирует транспорт веществ, а также имеет немалую роль в образовании спор.
Цитоплазма бактерий на 75 процентов произведена из воды. Состав цитоплазмы:
- Рыбосомы;
- мезосомы;
- аминокислоты;
- ферменты;
- пигменты;
- сахар;
- гранулы и включения;
- нуклеоид.
Обмен веществ у прокариотов возможен, как с участием кислорода, так и без его него. Большая их часть питаются уже готовыми питательными веществами органического происхождения. Очень мало видов способны сами синтезировать органические вещества из неорганических. Это сине-зеленые бактерии и цианобактерии, которые отыграли немалую роль в формировании атмосферы и насыщении её кислородом.
Размножение
В условиях, благоприятных для размножения, оно осуществляется почкованием или вегетативно. Бесполое размножение происходит в такой последовательности:
- Клетка бактерии достигает максимального объема и содержит необходимый запас питательных веществ.
- Клетка удлиняется, посередине появляется перегородка.
- Внутри клетки происходит дележ нуклеотида.
- ДНК основная и отделенная расходятся.
- Клетка делится пополам.
- Остаточное формирование дочерних клеток.
При таком способе размножения нету обмена генетической информацией, поэтому все дочерние клетки будут точной копией материнской.
Процесс размножения бактерий в неблагоприятных условиях более интересен. О способности полового размножения бактерий ученые узнали сравнительно недавно – в 1946 году. У бактерий нет разделения на женские и половые клетки. Но ДНК у них встречается разнополое. Две такие клетки при приближении друг к другу образовывают канал для передачи ДНК, происходит обмен участками – рекомбинация. Процесс довольно длительный, результатом которого являются две совершенно новые особи.
Большинство бактерий очень сложно увидеть под микроскопом, так как они не имеют своей окраски. Немногие разновидности имеют пурпурный или зеленый окрас, благодаря содержанию в них бактериохлорофилла и бактериопурпурина. Хотя если рассматривать некоторые колонии бактерий, становится ясно, что они выделяют окрашиваемые вещества в среду обитания и приобретают яркую окраску. Для того, чтобы подробней изучать прокариотов, их окрашивают.

Классификация
Классификация бактерий может быть основана на таких показателях, как:
- Форма
- способ передвижения;
- способ получения энергии;
- продукты жизнедеятельности;
- степень опасности.

Бактерии симбионты живут в содружестве с иными организмами.
Бактерии сапрофиты проживают на уже отмерших организмах, продуктах и органических отходах. Они способствуют процессам гниения и брожения.
Гниение очищает природу от трупов и других отходов органического происхождения. Без процесса гниения не было бы круговорота веществ в природе. Так в чем же состоит роль бактерий в круговороте веществ?
Бактерии гниения - это помощник в процессе расщепления белковых соединений, а также жиров и других соединений, содержащих в себе азот. Проведя сложную химическую реакцию, они разрывают связи между молекулами органических организмов и захватывают молекулы белка, аминокислот. Расщепляясь, молекулы высвобождают аммиак, сероводород и другие вредные вещества. Они ядовиты и могут вызывать отравление у людей и животных.
Бактерии гниения быстро размножаются в благоприятных для них условиях. Так как это не только полезные бактерии, но и вредные, то чтобы не допустить преждевременного гниения у продуктов, люди научились их обрабатывать: сушить, мариновать, солить, коптить. Все эти способы обработки убивают бактерии и не дают им размножаться.
Бактерии брожения при помощи ферментов способны расщеплять углеводы. Эту способность люди заметили еще в древние времена и используют такие бактерии для изготовления молочнокислых продуктов, уксусов, а также других продуктов питания до сих пор.
Бактерии, трудясь в совокупности с другими организмами, делают очень важную химическую работу. Очень важно знать какие есть виды бактерий и какую пользу или вред приносят для природы.
Значение в природе и для человека
Выше уже отмечалось большое значение многих видов бактерий (при процессах гниения и различных типах брожения), т.е. выполнение санитарной роли на Земле.
Бактерии также играют огромную роль в круговороте углерода, кислорода, водорода, азота, фосфора, серы, кальция и других элементов. Многие виды бактерий способствуют активной фиксации атмосферного азота и переводят его в органическую форму, способствуя повышению плодородия почв. Особо важное значение имеют те бактерии, которые разлагают целлюлозу, являющиеся основным источником углерода для жизнедеятельности почвенных микроорганизмов.
Сульфатредуцирующие бактерии участвуют в образовании нефти и сероводорода в лечебных грязях, почвах и морях. Так, насыщенный сероводородом слой воды в Черном море является результатом жизнедеятельности сульфатредуцирующих бактерий. Деятельность этих бактерий в почвах приводит к образованию соды и содового засоления почвы. Сульфатредуцирующие бактерии переводят питательные вещества в почвах рисовых плантаций в такую форму, которая становится доступной для корней этой культуры. Эти бактерии могут вызывать коррозию металлических подземных и подводных сооружений.
Благодаря жизнедеятельности бактерий почва освобождается от многих продуктов и вредных организмов и насыщается ценными питательными веществами. Бактерицидные препараты успешно используются для борьбы с многими видами насекомых-вредителей (кукурузным мотыльком и др.).
Многие виды бактерий используются в различных отраслях промышленности для получения ацетона, этилового и бутилового спиртов, уксусной кислоты, ферментов, гормонов, витаминов, антибиотиков, белково-витаминных препаратов и т.д.
Без бактерий невозможны процессы при дублении кожи, сушке листьев табака, выработке шелка, каучука, обработке какао, кофе, мочении конопли, льна и других лубоволокнистых растений, квашении капусты, очистке сточных вод, выщелачивании металлов и т.д.
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки | 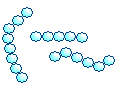 | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Существуют два типа систематики биологических объектов: филогенетическая, или естественная , в основе которой лежит установление родственных (генетических, эволюционных) связей между организмами, и практическая, или искусственная , целью которой является выявление степени сходства между организмами для быстрой их идентификации и установления принадлежности к определенным таксонам.
Большинство классификаций бактерий является искусственными. Они предназначены для определения той или иной группы микроорганизмов, представляющих интерес для исследователей
Классификация всех живых существ основана почти полностью на морфологических признаках организмов.
Морфология микроорганизмов изучает форму и особенности строения клеток, способы размножения и передвижения, и др. Морфологические признаки играют большую роль в идентификации микроорганизмов и их классификации.
У бактерий классификация имеет специфические особенности вследствие немногочисленности их морфологических признаков. Современная микробиология для классификации использует комплекс признаков: морфологические (форма клеток, наличие и характер расположения жгутиков, способ размножения, окраска по Граму, способность к образованию эндоспор; физиологические особенности (способ питания, получение энергии, состав продуктов обмена, отношение к воздействию температуры, рН, кислорода и др. факторам) ; культуральные (характер роста на различных питательных средах культуры бактерии; на жидких средах - наличие пленки, мути, осадка; на плотных средах - тип колоний и их особенности).
В настоящее время большое значение имеют биохимические (генотипические) признаки, т.е. особенности нуклеотидного состава ДНК. Достоверно известно, что особи одного и того же вида имеют одинаковый состав оснований ДНК, а у видов, принадлежащих к одному роду, нуклеотидный состав имеет близкие значения. По совокупности морфологических, физиологических, культуральных и биохимических признаков бактерии могут быть отнесены к тому или иному виду.
В последние годы получила признание искусственная классификация бактерий, предложенная Р.Мюрреем (R.Murray) в 1978 году. Согласно этой классификации царство прокариот «Procaryotae» подразделено на четыре отдела. Распределение микроорганизмов по отделам основано главным образом, на наличии или отсутствии клеточных стенок и особенностей их строения. Для микробиологии пищевых производств имеют значения два отдела:
В первый отдел Firmicutes («Firmus» - толстый, основательный) или «толстокожие», отнесены все бактерии, для которых характерно строение клеточной стенки по типу Грам+ бактерий: все кокки, молочнокислые бактерии (педиококки - Pediococcus, лактобациллы – Lactobacillus, стреп-тококки – Streptococcus и лейконосток – Leuconostoc), палочковидные спорообразующие бактерии (Bacillus, Clostridium) и актиномицеты. Второй отдел Gracilicutes («Gracilus» - тонкий, изящный, «cutes» - кожа) или «тонкокожие», объединяет все бактерии, которые имеют клеточную стенку, характерную для Грам- бактерий: род Pseudomonas (некоторые гнилостные бактерии и др.), роды Acetobacter и Gluconobacter (уксуснокислые бактерии), используемые в производстве уксуса, а также вредители бродильных производств. К Грам- палочкам относится и многочисленная группа – энтеробактерии (бактерии кишечной группы), в т.ч. и род Escherichia. Некоторые из бактерий кишечной группы постоянно населяют кишечник человека и животных. Другие - являются возбудителями инфекционных желудочно-кишечных заболеваний (дизентерии, брюшного тифа, паратифов), передающихся через пищевых продукты, и пищевых отравлений.
Контрольные вопросы:
1. Каковы основные формы бактерий пищевых производств? 2. Назовите основные функции и химический состав клеточной стенки бактерий? 3. Какие фукнции в бактериальной клетке выполняет цитоплазматическая мембрана? 4. Чем представлен генетический аппарат у прокариот? 5.Что такое плазмиды, у каких бактерийони присутствуюти какие функции выполняют? 6.Как передвигаются бактерии? 7.Какие функции выполняют эндоспоры у бактерий, и при каких условиях они образуются? 8.Назовите основной принцип классификации бактерий?
Эукариоты - микроскопические грибы (мицелиальные грибы и дрожжи)
Для мицелиальных грибов характерны разнообразные способы и органы размножения. Различия в строении мицелия и способах размножения используются для классификации грибов. Клетки грибов имеют ветвящиеся нити - гифы с верхушечным ростом и боковым ветвлением, переплетаясь они образуют мицелий (грибницу).
Грибы размножаются вегетативным, бесполым и половым путями.
Вегетативное размножение осуществляется отдельными участками мицелия, т.е. без образования специализированных органов размножения.
При бесполом и половом размножении образуются специализированные клетки - споры, с помощью которых и осуществляется размножение.
Образованию спор при бесполом размножении предшествует митотическое деление ядра , при котором образуются два дочерних ядра с набором хромосом, идентичным набору родительской клетки.
Споры при бесполом размножении образуются на особых плодо-
носящих гифах воздушного мицелия, внешне отличающихся от вегета-
тивных гиф.
У низших грибов споры образуются внутри специальных клеток -спорангиев, они носят название спорангиоспоры . У высших грибов споры образуются экзогенно (наружно) на гифах воздушного мицелия и носят название конидии .
Рис.12. Органы вегетативного и бесполого размножения грибов: а - оидии; б - хламидоспоры; в - спорангиоспоры; г – конидии
Образованию спор при половом размножении, предшествует слияние (копуляция ) двух половых клеток – гамет и их ядер. Образуется диплоидная клетка - зигота , содержащая двойной набор хромосом. Затем следует процесс редукционного деления - мейоз, сопровождающийся перераспределением отцовских и материнских признаков, приводящий к уменьшению числа хромосом до исходного и увеличению разнообразия видов. В результате образуются специализированные органы размножения. Развитие этих органов, формы полового процесса у грибов многообразны.
Классификация грибов . Подразделение грибов на классы основано на использовании комплекса признаков, ведущими из которых являются особенность состава клеточной стенки, типы полового и бесполого размножения. Согласно современной классификации все грибы распределены по следующим классам:
Класс Chytridiomycetes (Хитриодиомицеты )
Synchytrium - является возбудителем рака картофеля.
Класс Zygomycetes (Зигомицеты ): Род Мucor - вызывают порчу пищевых продуктов, образуя пушистые налеты.


Рис.13. Род Мucor Рис.14. Род Rhizopus
Грибы рода Rhizopus вызывают так называемую «мягкую - гниль» ягод, плодов и овощей. Мукоровые грибы образуют органические кислоты и ферменты, способны вызывать слабое спиртовое брожение.
Класс Ascomycetes (Аскомицеты ): К аскомицетам относятся имеющие большое значение аспергилловые и пеницилловые грибы.



Рис.15. Aspergillus niger Рис.16. Penicillium chrysogenum
Сумчатые грибы широко распространены в природе. Многие из них являются возбудителями порчи плодов и овощей, (особенно при их хранении - различные гнили), многих пищевых продуктов. Некоторые из них вызывают повреждение промышленных изделий и материалов (текстиля, резины, целлофана, пластмасс и т.д.). Отдельные представители аспергилловых и пеницилловых грибов используются в промышленности. Так некоторые пенициллы являются продуцентами антибиотиков – пенициллина, цефалоспорина, гризеофульвина, цитринина и др. Penicillium roqueforti , Penicillium camemberti используют в производстве сыра сортов Рокфор и Камамбер; Aspergillus niger – для промышленного получения лимонной кислоты; A. oryzae, A. awamori - для получения ферментных препаратов. Некоторых аспергиллы являются патогенными для человека и животных, вызывая поражение дыхательных путей (отомикозы, аспергиллез и эмфизему легких), кожи (дерматомикозы), слизистой рта.
В последние полвека особое внимание ученых обращено на вторичные метаболиты мицелиальных грибов, развивающихся на пищевом сырье растительного и животного происхождения и на пищевых продуктах и кормах, - микотоксины . Примерно 60 – 75% грибов-возбудителей порчи пищевых продуктов и кормов животных являются токсичными и высокотоксичными. Употребление заплесневелых пищевых продуктов крайне опасно для здоровья человека и животных. Многочисленными исследованиями установлено гепатотропное, канцерогенное и мутагенное действие на организм человека и животных афлатоксинов, охратоксинов, патулина, рубратоксина и др., выделяемых грибами Aspergillus flavus, A. ochraceus, Penicillium veridatum, P.islandicum, P. rubrum, P. expansum и др. Все микотоксины опасны даже в незначительных количествах и с трудом поддаются деградации (разрушению).


Рис.17. Claviceps (спорынья) Рис.18. Monilia (монилия)
К плодосумчатым аскомицетам относятся также трюфели и сморчки, плодовые тела которых употребляют в пищу, а также строчки, считающиеся условно съедобными, так как некоторые их виды ядовиты.


Рис.19: а – шапочка сморчковая; б - строчок осенний.

Рис.20. Трутовый гриб
Эта способность у них выражена гораздо резче, чем у высших растений, лишайников и других организмов. Вот почему нельзя собирать грибы в местах загрязненных отходами производства. Накопление указанных элементов вызывает ряд необратимых перестроек в биохимическом аппарате грибов. Это явление пока что мало изучено и поэтому представляет угрозу для здоровья человека.
Класс Deuteromycetes (Дейтеромицеты ): Половое размножение у них отсутствует, размножаются они только бесполым путем, в основном конидиями, которые, как и конидиеносцы, имеют самую различную форму, вид и окраску. У некоторых видов не образуется специализированных органов размножения, и они размножаются кусочками мицелия.



Рис.21. Род Fusarium (Фузариум) Рис.22. Род Botrytis
Род Botrytis гриб вызывает кагатную гниль сахарной свеклы; развиваясь на винограде, плодах и ягодах, размягчает гкани, которые становятся водянистыми. Продуцирует ферменты пектиназу, цеплюлазу, инвертазу др.
Виды Alternaria широко распространены в почве и на растительных остатках. Гриб вызывает заболевание многих сельскохозяйственных растений альтернариоз. На пищевых продуктах образуют черные вдавленные пятна (черная сухая гниль моркови, черная пятнистость капусты). Когда пораженные участки листа выпадают, образуются дырочки.
Род Geotrichum развивается на поверхности кисломолочных продуктов, сыров, квашеных овощей, прессованных дрожжей, стенках оборудования, сырых помещений. Некоторые виды рода Geotrichum вызывают порчу плохо высушенного хмеля.
Род Monilia являются активными возбудителями порчи плодов, которые превращаются в так называемые «мумии». Представители этого рода, относящиеся к классу дейтеромицетов, существуют в конидиальной стадии.
Род Cladosporium. Грибы нередко обнаруживаются при холодильном хранении на различных пищевых продуктах в виде бархатистых темно-оливковых (до черного цвета) пятен.
Род Helminthosporium Болезни злаков, вызываемые грибами этого рода, называют гельминтоспориозами. Некоторые виды этого рода - сапрофиты и развиваются на корнях, листьях, сухих ветвях, стеблях, стеблях древесины и травянистых растений.

Рис. 23. Род Helminthosporium
Дрожжи являются одноклеточными грибными неподвижным организмами, не имеющими настоящего мицелия. Обитают в основном на растениях, где имеются сахаристые вещества, которые они сбраживают (нектар цветов, сочные фрукты, ягоды, особенно перезрелые и поврежденные, листья, стволы березы во время сокотечения и дуба во время слизетечения, почва). Клетки дрожжей имеют овальную, цилиндрическую, яйцевидную, лимоновидную, колбовидную, треугольную, стреловидную и серповидную форму. Некоторые виды дрожжей наряду с круглыми и овальными клетками могут образовывать удлиненные, а также псевдомицелий. Дрожжевые клетки, значительно крупнее бактериальных.
Как и все грибы, дрожжи являются неподвижными организмами. Дрожжи имеют достаточно сложную структурную организацию, типичную для эукариотных организмов.
Дрожжи размножаются вегетативно и спорами, образующимися бесполым и половым путем. Способ размножения является важным признаком для классификации дрожжей. Наиболее распространенным способом вегетативного размножения является почкование.
Если при почковании вновь возникающие клетки не отделяются друг от друга, то образуется псевдомицелий. Размножение делением, характерное для дрожжей цилиндрической формы, встречается реже. У дрожжей лимоновидной формы наблюдается так называемое почкующееся деление, при котором на широком основании формируется почка, процесс заканчивается появлением хорошо заметной септы в районе перешейка.
При половом размножении их появлению предшествует слияние клеток и последующее объединение ядер, при бесполом размножении предварительное слияние клеток и ядер не происходит. Половое размножение большинства дрожжей связано с образованием асков (сумок) и аскоспор.
Образованию аскоспор предшествует копуляция (слияние содержимого двух клеток и их ядер). Образуется зигота, в которой затем формируются споры: ядро делится мейозом, вокруг новых ядер уплотняется цитоплазма, и они покрываются плотной оболочкой. Такие дрожжи относятся к классу аскомицетов. Аскоспоры могут образовывать только молодые клетки на полноценной питательной среде и перенесенные в условия голодания, плохого снабжения кислородом и влагой. У различных видов дрожжей в аске образуется
2 - 4, а иногда 8 спор. При спорообразовании замедлен обмен веществ и жизнедеятельность клеток. Такое состояние обеспечивает их выживаемость в условиях, неблагоприятных для вегетативного размножения.
Аскоспоры устойчивы к действию высокой температуры, высушиванию, но они менее термостабильны, чем бактериальные споры, и погибают при температуре 60°С. При условиях, благоприятных для вегетативного развития, на свежей питательной среде споры прорастают и снова размножаются вегетативно. Поскольку дрожжи по существу являются одноклеточными немицелиальными грибами, они включены в классификацию грибов. Они распределены по трем классам грибов –Ascomycetes, Basidiomycetes и Deuteromycetes .
Аскомицетовые дрожжи включают примерно 2/3 дрожжей. Среди них наибольшее практическое значение имеют сахаромицеты, объединяющие более половины известных родов дрожжей. Особо важная роль принадлежит Saccharomyces cerevisiae (крупные овальные клетки) в производстве этилового спирта, пива, кваса и в хлебопечении и Saccharomyces ellpsoideus (крупные эллиптические клетки) - их используют преимущественно в виноделии.
Рис.25. Saccharomyces cerevisiae
Дрожжи класса Deuteromycetes наибольшее значение имеют роды Candida, Torulopsis и Rhodotorula. Candida имеют удлиненную форму клеток, сочетания которых образуют примитивный псевдомицелий. Многие из них не вызывают спиртовое брожение и являются вредителями в бродильных производствах (например, Candida mycoderma ). Другие представители рода Candida являются вредителями в дрожжевом производстве, снижают качество хлебопекарных дрожжей, так как относятся к слабосбраживающим видам. Некоторые из них вызывают порчу квашенных овощей, безалкогольных и ряда других напитков и продуктов. Среди этих дрожжей имеются патогенные виды, вызывающие кандидозы, поражающие слизистые оболочки ротовой полости, носоглотки и других органов человека. Различные виды дрожжей рода Candida используются для получения кормового белка и белково-витаминных концентратов (БВК).
Дрожжи рода Torulopsis способны вызывать слабое спиртовое брожение и используются в производстве кефира и кумыса. Некоторые применяются для промышленного получения кормового белка.
Дрожжи рода Rhodotorula используются для промышленного получения кормовых белково-витаминных концентратов, которые служат источником жирорастворимого витамина А для животных. Другие представители этого рода накапливают в клетках много липидов и используются в микробиологической промышленности как продуценты липидов.
Вирусы

Рис.26. Бактериофаг: А – модель фага; Б – фаг, инъецкцировавший свою ДНК в клетку
В медицине бактериофаги применяются для лечения некоторых заболеваний, например дизентерии.
Контрольные вопросы :
1 . Каковы морфологические особенности и способы размножения мицелиальных грибов? 2. Каковы особенности строения и размножения дрожжей? 3. Расскажите об основных принципах классификации прокариот и эукариот. 4. Назовите основных представителей отдельных классов эукариот и их практическое значение.5. Расскажите остроении и практическом значении вирусов и фагов.
Обмен веществ у микроорганизмов отличается чрезвычайным разнообразием. Это связано со способностью микроорганизмов использовать для обмена веществ широкий круг органических и минеральных соединений. Такая способность обусловливается наличием у микроорганизмов большого разнообразия ферментов. На активность ферментов влияют температура, рН и другие факторы внешней среды - воздействие химических веществ среды, лучистая энергия и др. Физиологические процессы, протекающие в клетках микроорганизмов, почти полностью зависят от активности ферментов, поэтому любой фактор, действующий на фермент, будет воздействовать и на метаболизм микроорганизмов.
Каждому виду микроорганизмов свойственен определенный набор ферментов, постоянно присутствующих в клетке (т.н. конститутивные ферменты). В то же время некоторые ферменты синтезируются клеткой только тогда, когда в среде появляется соответствующий субстрат. Такие ферменты называют индуктивными.
По характеру действия ферменты подразделяются на экзоферменты, выделяемые клеткой в окружающую среду, и эндоферменты. прочно связанные с клеточными структурами (митохондриями, цитоплазматической мембраной и мезосомами) и действуют внутри клетки. И те, и другие играют важную роль в обмене веществ микроорганизмов. Экзоферменты (обычно гидролазы) катализируют реакции вне клетки. К эндоферментам относятся оксидоредуктазы (окислительно-восстановительные ферменты), трансферазы (ферменты переноса) и др., играющие важную роль в энергетическом обмене.
Конструктивный обмен веществ заключается в биосинтезе основных клеточных компонентов из поступивших в клетку веществ питательной среды. Конструктивный обмен направлен на синтез четырех основных типов биополимеров: белков, нуклеиновых кислот, полисахаридов и липидов. Синтез протекает как серия последовательных реакций с образованием разнообразных промежуточных продуктов метаболизма. Кроме того, уровни развития биосинтетических способностей микроорганизмов различны. Именно поэтому микроорганизмы резко отличаются друг от друга по своим потребностям в питательных веществах. Независимо от их потребностей в питательной среде должны содержаться все элементы, которые имеются в клетках микроорганизмов. По отношению к источникам углерода все микроорганизмы делятся на две большие группы: автотрофы и гетеротрофы . Соответственно и тип питания этих микроорганизмов называется либо автотрофным, либо гетеротрофным. Микроорганизмы, использующие для биосинтеза веществ клетки неорганический источник углерода (СО 2),называются автотрофами. Микроорганизмы, которые не могут использовать СО 2 в качестве единственного источника углерода, и нуждаются в органических соединениях, называются гетеротрофами. К гетеротрофам относится большинство микроорганизмов.
Многие гетеротрофные микроорганизмы для синтеза веществ клетки в качестве источника углерода используют в основном углеводы, а также спирты, но, кроме того, могут использовать липиды, белки, аминокислоты (их углеродный скелет) и гораздо реже - органические кислоты. По отношению к источнику азота микроорганизмы делятся на аминоавтотрофы и аминогетеротрофы. Аминоавтотрофы усваивают азот из минеральных соединений (нитратов, нитритов, аммонийных солей и т.п.) Аминогетеротрофы нуждаются в готовых органических азотсодержащих соединениях (белках, аминокислотах, пуринах, пиримидинах), которые они используют одновременно как источник углерода и азота.
Сапрофиты питаются за счет органических веществ отмерших животных и растений. К ним относятся гнилостные бактерии, мицелиальные грибы, актиномицеты, дрожжи, бактерии-возбудители процессов брожения и др.
Поступление воды и питательных веществ из окружающей среды и выделение продуктов метаболизма у микроорганизмов происходит через всю поверхность клеток. Вещества питательной среды должны обладать растворимостью в воде или в липидах, поскольку они могут проникать внутрь микробной клетки только в растворенном виде; продукты метаболизма выводятся из клетки также в растворенном состоянии. Нерастворимые сложные органические вещества (белки, полисахариды, жиры и др.) питательной среды предварительно подвергаются расщеплению вне клетки на более низкомолекулярные соединения, обладающие растворимостью в воде (аминокислоты, моносахариды, органические кислоты и др.), с помощью выделяемых во внешнюю среду микроорганизмами гидролитических ферментов.
Молекулы воды, некоторых газов О 2 , Н 2 , N 2 , некоторые ионы, концентрация которых во внешней среде выше, чем в клетке, перемещаются через ЦПМ внутрь клетки путем пассивной диффузии. Пассивный перенос веществ протекает до тех пор, пока концентрация веществ по обе стороны ЦПМ не выровняется. Вода - основное вещество, которое проникает в клетку путем пассивной диффузии.
В клетку из питательной среды поступают только те питательные вещества, для которых в ЦПМ имеются соответствующие переносчики, и в этом проявляется избирательная проницаемость ЦПМ.
Пермеазы обладают строгой специфичностью к субстрату, т.е. каждый из них переносит только определенное вещество. Переносчик вступает во взаимодействие с веществом на наружной стороне ЦПМ, и этот комплекс диффундирует через ЦПМ к внутренней стороне ЦПМ, комплекс распадается и затем вещество передается в цитоплазму. После этого переносчики "захватывают" определенные продукты обмена, выносят их из клетки и процесс повторяется. Таким образом, в клетку из питательной среды поступают только те вещества, для которых в ЦПМ имеются соответствующие переносчики, и в этом проявляется избирательная проницаемость ЦПМ.
С помощью переносчиков осуществляется перенос растворенных веществ питательной среды путем облегченной диффузии и активного транспорта.
Облегченная диффузия происходит по градиенту концентрации, как и пассивная диффузия, она протекает тоже без затраты энергии, но с большей скоростью.

Рис 27. .Транспорт веществ через цитоплазматическую мембрану:
а - цитоплазма: б - мембрана; в- окружающая среда: р – переносчик
Активный транспорт веществ идет против градиента концентрации, т.е. от меньшей концентрации к большей, что обязательно сопровождается затратой энергии. Попав внутрь клетки, вещество освобождается от переносчика также с затратой энергии. При активном транспорте скорость поступления вещества в клетку достигает максимума уже при малой концентрации его в питательной среде, причем концентрация этого вещества в клетке может значительно превысить его концентрацию в питательной среде.
Прокариоты и эукариоты различаются по механизмам транспорта - у прокариот избирательное поступление питательных веществ происходит путем активного транспорта, у эукариот - путем облегченной диффузии. Вывод продуктов обмена из клеток микроорганизмов чаще всего осуществляется путем облегченной диффузии.
2.2. Классификация и морфология бактерий
Классификация бактерий . Решением Международного кодекса для бактерий рекомендованы следующие таксономические категории: класс, отдел, порядок, семейство, род, вид. Название вида соответствует бинарной номенклатуре, т. е. состоит из двух слов. Например, возбудитель сифилиса пишется как Treponema pallidum . Первое слово - на-
звание рода и пишется с прописной буквы, второе слово обозначает вид и пишется со строчной буквы. При повторном упоминании вида родовое название сокращается до начальной буквы, например: Т. pallidum .
Бактерии относятся к прокариотам, т.е. доядерным организмам, поскольку у них имеется примитивное ядро без оболочки, ядрышка, гистонов. а в цитоплазме отсутствуют высокоорганизованные органеллы (митохондрии, аппарат Гольджи, лизосомы и др.)
В старом Руководстве Берджи по систематической бактериологии бактерии делили по особенностям клеточной стенки бактерий на 4 отдела:Gracilicutes - эубактерии с тонкой клеточнойстенкой, грамотрицательные; Firmicutes - эубактерии с толстой клеточной стенкой, грамположи-тельные; Tenericutes - эубактерии без клеточной стенки; Mendosicutes - архебактерии с дефектной клеточной стенкой.
Каждый отдел был разделен на секции, или группы, по окраске по Граму, форме клеток, потребности в кислороде, подвижности, особенностям метаболизма и питания.
Согласно 2-му изданию (2001 г.) Руководства Берджи, бактерии делят на 2 домена: «Bacteria» и «Archaea» (табл. 2.1).
Таблица. Характеристика доменов Bacteria и Archaea
|
Домен «Bacteria» (эубактерии) |
Домен «Archae а» (архебактерии) |
|
В домене «Bacteria» можно выделить следующие бактерии: 1)бактерии с тонкой клеточной стенкой, грамотрицательные*; 2)бактерии с толстой клеточной стенкой, грамположительные**; 3)бактерии бет клеточной стенки (класс Mollicutes - микоплаз- мы) |
Архсбактерии не содержат пепти-догликан в клеточной стенке. Они имеют особые рибосомы и рибосом-ные РНК (рРНК). Термин «архебактерии- появился в 1977 г. Это одна из древних форм жизни, на что указывает приставка «архе». Среди них нет возбудителей инфекций |
*Среди тонкостенных грамотрицательных эубактерий различают:
сферические формы, или кокки (гонококки, менингококки, вейлонеллы);
извитые формы - спирохеты и спириллы;
палочковидные формы, включая риккетсии.
** К толстостенным грамположительным эубактериям относят:
сферические формы, или кокки (стафилококки, стрептококки, пневмококки);
палочковидные формы, а также актиномицеты (ветвящиеся, нитевидные бактерии), коринебактерии (булавовидные бактерии), микобактерии и бифидобактерии (рис. 2.1).
Большинство грамотрицательных бактерий объединены в тип протеобактериий. основанный на сходстве по рибосомной РНК «Proteobacteria» - по имени греческого бога Протеуса. принимавшего разнообразные облики). Они появились от общего фотосинте-тического предка.
Грамположительные бактерии, согласно изученным последовательностям рибосомной РНК, являются отдельной филогенетической группой с двумя большими подотделами - с высоким и низким соотношением G + C (генетическое сходство). Как и протеобактерии, эта группа метаболически разнообразная.
В домен « Bacteria » входят 22 типа, из кото рых медицинское значение имеют следующие:
Тип Proteobacteria
Класс Alphaproteobacteria. Роды : Rickettsia, Orientia, Ehrlichia, Bartonella, Brucella
Класс Betaproteobacteria. Роды : Burkholderia, Alcaligenes, Bordetella, Neisseria, Kingella, Spirillum
Класс Gammaproteobacteria. Роды : Francisella, Legionella, Coxiella, Pseudomonas, Moraxella, Acinetobacter, Vibrio, Enterobacter, Callimatobacterium, Citrobacter, Edwardsiella, Erwinia, Escherichia, Hafnia, Klebsiella, Morganella, Proteus, Providencia, Salmonella, Serratia, Shigella, Yersinia, Pasteurella
Класс Deltaproteobacteria. Род: Bilophila
Класс Epsilonproteobacteria. Роды : Campylobacter, Helicobacter, Wolinella
Тип Firmicutes (главным образом грамполо жительные )
Класс Clostridia. Роды : Clostridium, Sarcina, Peptostreptococcus, Eubacterium, Peptococcus, Veillonella (грамотрицательные)
Класс Mollicutes. Роды: Mycoplasma, Ureaplasma
Класс Bacilli. Роды : Bacillus, Sporosarcina, Listeria, Staphylococcus, Gemella, Lactobacillus, Pediococcus, Aerococcus, Leuconostoc, Streptococcus, Lactococcus
Тип Actinobacteria
Класс Actinobacteria. Роды : Actinomyces, Arcanodacterium, Mobiluncus, Micrococcus, Rothia, Stomatococcus, Corynebacterium, Mycobacterium, Nocardia, Propionibacterium, Bifidobacterium, Gardnerella
Тип Clamydiae
Класс Clamydiae. Роды : Clamydia, Clamydophila
Тип Spirochaetes
Класс Spirochaetes. Роды : Spirochaeta, Borrelia, Treponema, Leptospira
Тип Bacteroidetes
Класс Bacteroidetes. Роды : Bacteroides, Porphyromonas, Prevotella
Класс Flavobacteria. Роды: Flavobacterium
Подразделение бактерий по особенностям строения клеточной стенки связано с возможной вариабельностью их окраски в тот или иной цвет по методу Грама. Согласно этому методу, предложенному в 1884 г. датским ученым X. Грамом, в зависимости от результатов окраски бактерии делятся на грамположительные, окрашиваемые в сине-фиолетовый цвет, и грамотрицательные, красящиеся в красный цвет. Однако оказалось, что бактерии с так называемым грамположи-тельным типом клеточной стенки (более толстой, чем у грамотрицательных бактерий), например, бактерии рода Mobiluncus и некоторые спорообразующие бактерии, вместо обычной грамположительной окраски имеют грамотрицательную окраску. Поэтому для таксономии бактерий бульшую значимость, чем окраска по Граму, имеют особенности строения и химического состава клеточных стенок.
2.2.1. Формы бактерий
Различают несколько основных форм бактерий (см. рис. 2.1) - кокковидные, палочковидные, извитые и ветвящиеся, нитевидные формы бактерий.
Сферические формы, или кокки, - шаровидные бактерии размером 0,5-1,0 мкм*, которые по взаимному расположению делятся на микрококки, диплококки, стрептококки, тетракокки, сарцины и стафилококки.
Микрококки (от греч. micros - малый) - отдельно расположенные клетки.
Диплококки (от греч. diploos - двойной), или парные кокки, располагаются парами (пневмококк, гонококк, менингококк), так как клетки после деления не расходятся. Пневмококк (возбудитель пневмонии) имеет с противоположных сторон ланцетовидную форму, а гонококк (возбудитель гонореи) и менингококк (возбудитель эпидемического менингита) имеют форму кофейных зерен, обращенных вогнутой поверхностью друг к другу.
Стрептококки (от греч. streptos - цепочка) - клетки округлой или вытянутой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления.
Сарцины (от лат. sarcina - связка, тюк) располагаются в виде пакетов из 8 и более кокков, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях.
Стафилококки (от греч. staphyle - виноградная гроздь) - кокки, расположенные в виде грозди винограда в результате деления в разных плоскостях.
Палочковидные бактерии различаются по размерам, форме концов клетки и взаимному расположению клеток. Длина клеток варьирует от 1,0 до 10 мкм, толщина - от 0,5 до 2,0 мкм. Палочки могут быть правильной (кишечная палочка и др.) и неправильной (коринебактерии и др.) формы, в том числе ветвящиеся, например, у актиномицетов. К наиболее мелким палочковидным бактериям относятся риккетсии.
Концы палочек могут быть как бы обрезанными (сибиреязвенная бацилла), закругленными (кишечная палочка), заостренными (фузобактерии) или в виде утолщения. В последнем случае палочка похожа на булаву (коринебактерии дифтерии).
Слегка изогнутые палочки называются вибрионами (холерный вибрион). Большинство палочковидных бактерий располагается беспорядочно, так как после деления клетки расходятся. Если после деления клетки остаются связанны-
ми общими фрагментами клеточной стенки и не расходятся, то они располагаются под углом друг к другу (коринебактерии дифтерии) или образуют цепочку (сибиреязвенная бацилла).
Извитые формы - спиралевидные бактерии, например спириллы, имеющие вид штопоро-образно извитых клеток. К патогенным спириллам относится возбудитель содоку (болезнь укуса крыс). К извитым также относятся кам-пилобактерии и хеликобактерии, имеющие изгибы как у крыла летящей чайки; близки к ним и такие бактерии, как спирохеты. Спирохеты - тонкие, длинные, извитые
спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной сгибательными изменениями клеток. Спирохеты состоят из наружной мембраны
клеточной стенки), окружающей протоплазматический цилиндр с цитоплазматической мембраной и аксиальной нитью (аксистиль). Ахсиальная нить находится под наружной мембраной клеточной стенки (в периплазме) и как бы закручивается вокруг протоплазма-тического цилиндра спирохеты, придавая ей винтообразную форму (первичные завитки спирохет). Аксиальная нить состоит из перип-лазматических фибрилл - аналогов жгутиков бактерий и представляет собой сократительный белок флагеллин. Фибриллы прикреплены к концам клетки (рис. 2.2) и направлены навстречу друг другу. Другой конец фибрилл свободен. Число и расположение фибрилл варьируют у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками. Спирохеты
плохо воспринимают красители. Обычно их окрашивают по Романовскому-Гимзе или серебрением. В живом виде спирохеты исследуют с помощью фазово-контрастной или темнопольной микроскопии.
Спирохеты представлены 3 родами, патогенными для человека: Treponema , Borrelia , Leptospira .
Трепонемы (род Treponema) имеют вид тонких штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками. Вокруг протопласта трепонем расположены 3-4 фибриллы (жгутики). В цитоплазме имеются цитоплазматические филаменты. Патогенными представителями являются Т. pallidum - возбудитель сифилиса, Т. pertenue - возбудитель тропической болезни - фрам-безии. Имеются и сапрофиты - обитатели полости рта человека, ила водоемов.
Боррелии (род Borrelia ), в отличие от трепонем, более длинные, имеют по 3-8 крупных завитков и 7-20 фибрилл. К ним относятся возбудитель возвратного тифа (В. recurrentis ) и возбудители болезни Лайма (В. burgdorferi и др.).
Лептоспиры (род Leptospira ) имеют завитки неглубокие и частые - в виде закрученной веревки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв S или С; имеют 2 осевые нити (жгутики). Патогенный представитель L . in terrogans вызывает лептоспироз при попадании в организм с водой или пищей, приводя к развитию кровоизлияний и желтухи.
нием в цитоплазме, а некоторые - в ядре инфицированных клеток. Обитают в членистоногих (вшах, блохах, клещах) которые являются их хозяевами или переносчиками. Свое название риккетсии получили по имени X. Т. Риккетса - американского ученого, впервые описавшего одного из возбудителей (пятнистая лихорадка Скалистых гор). Форма и размер риккетсии могут меняться (клетки неправильной формы, нитевидные) в зависимости от условий роста. Структура риккетсии не отличается от таковой грамотрицательных бактерий.
Риккетсии обладают независимым от клетки хозяина метаболизмом, однако, возможно, они получают от клетки хозяина макроэр-гические соединения для своего размножения. В мазках и тканях их окрашивают по Романовскому-Гимзе, по Маккиавелло- Здродовскому (риккетсии красного цвета, а инфицированные клетки - синего).
У человека риккетсии вызывают эпидемический сыпной тиф (Rickettsia prowazekii ), клещевой риккетсиоз (R . sibirica ), пятнистую лихорадку Скалистых гор (R . rickettsii ) и другие риккетсиозы.
Элементарные тельца попадают в эпителиальную клетку путем эндоцитоза с формированием внутриклеточной вакуоли. Внутри клеток они увеличиваются и превращаются в делящиеся ретикулярные тельца, образуя скопления в вакуолях (включения). Из ретикулярных телец образуются элементарные тельца, которые выходят из клеток путем эк-зоцитоза или лизиса клетки. Вышедшие из
клетки элементарные тельца вступают в новый цикл, инфицируя другие клетки (рис. 16.11.1). У человека хламидии вызывают поражения глаз (трахома, конъюнктивит), уро-генитального тракта, легких и др.
Актиномицеты - ветвящиеся, нитевидные или палочковидные грамположительные бактерии. Свое название (от греч. actis - луч, mykes - гриб) они получили в связи с образованием в пораженных тканях друз - гранул из плотно переплетенных нитей в виде лучей, отходящих от центра и заканчивающихся кол-бовидными утолщениями. Актиномицеты, как и грибы, образуют мицелий - нитевидные переплетающиеся клетки (гифы). Они формируют субстратный мицелий, образующийся в результате врастания клеток в питательную среду, и воздушный, растущий на поверхности среды. Актиномицеты могут делиться путем фрагментации мицелия на клетки, похожие на палочковидные и кокко-видные бактерии. На воздушных гифах акти-номицетов образуются споры, служащие для размножения. Споры актиномицетов обычно не термостойки.
Общую филогенетическую ветвь с актино-мицетами образуют так называемые нокарди-оподобные (нокардиоформные) актиномицеты- собирательная группа палочковидных, неправильной формы бактерий. Их отдельные представители образуют ветвящиеся формы. К ним относят бактерии родов Corynebacterium , Mycobacterium , Nocardianjxp . Нокардиоподобные актиномицеты отличаются наличием в клеточной стенке Сахаров арабинозы, галактозы, а также миколовых кислот и больших количеств жирных кислот. Миколовые кислоты и липиды клеточных стенок обуславливают кис-лотоустойчивость бактерий, в частности ми-кобактерий туберкулеза и лепры (при окраске по Цилю-Нельсену они имеют красный цвет, а некислотоустойчивые бактерии и элементы ткани, мокроты - синий цвет).
Патогенные актиномицеты вызывают акти-номикоз, нокардии - нокардиоз, микобакте-рии - туберкулез и лепру, коринебактерии - дифтерию. Сапрофитные формы актиномицетов и нокардиеподобных актиномицетов широко распространены в почве, многие из них являются продуцентами антибиотиков.
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной «сдерживающая» высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов, имеет рецепторы для бактериофагов, бактериоцинов и различных веществ. Наиболее толстая клеточная стенка у грамположительных бактерий (рис. 2.4 и 2.5). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более.
Микоплазмы - мелкие бактерии (0,15-1,0 мкм), окруженные только цитоплазматической мембраной. Они относятся к классу Mollicutes , содержат стеролы. Из-за отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют разнообразную форму: кокковидную, нитевидную, колбовидную. Эти формы видны при фазово-кон-трастной микроскопии чистых культур микоплазм. На плотной питательной среде микоплазмы образуют колонии, напоминающие яичницу-глазунью: центральная непрозрачная часть, погруженная в среду, и просвечивающая периферия в виде круга.
Микоплазмы вызывают у человека атипичную пневмонию (Mycoplasma pneumoniae ) и поражения мочеполового тракта (М. homi - nis и др.). Микоплазмы вызывают заболевания не только у животных, но и у растений. Достаточно широко распространены и непатогенные представители.
2.2.2. Структура бактериальной клетки
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их улыратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и ядра, называемого нуклеоидом. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили (рис. 2.3). Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (му-реин, мукопептид), составляющий 40-90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного, с поперечными пептидными сшивками, пептидогликана.
Пептидогликан представлен параллельно расположенными молекулами гликана . состоящего из повторяющихся остатков N-аце-тилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эти связи разрывает лизоцим, являющийся аце-тилмурамидазой. Гликановые молекулы соединены через N-ацетилмурамовую кислоту поперечной пептидной связью из четырех аминокислот (тетрапептида ). Отсюда и название этого полимера - пептидогликан.
Основу пептидной связи пептидогликана грамотрицательных бактерий составляют тетрапеп-тиды, состоящие из чередующихся L- и D-ами-нокислот, например: L-аланин - D-глутаминовая кислота - мезо-диаминопимелиновая кислота - D-аланин. У Е. coli (грамотрицательная бактерия) пептидные цепи соединены друг с другом через D-аланин одной цепи и мезо-диаминопимели-
новую кислоту - другой. Состав и строение пептидной части пептидогликана грамотрицательных бактерий стабильны в отличие от пептидогликана грамположительных бактерий, аминокислоты которого могут отличаться по составу и последовательности. Тетрапептиды пептидогликана у грамположительных бактерий соединены друг с другом полипептидными цепочками из 5 остатков
глицина (пентаглицина). Вместо мезо-диамино-пимелиновой кислоты они часто содержат лизин. Элементы гликана (ацетилглюкозамин и аце-тилмурамовая кислота) и аминокислоты тетра-пептида (мезо-диаминопимелиновая и D-глу-таминовая кислоты, D-аланин) являются отличительной особенностью бактерий, поскольку отсутствуют у животных и человека.
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого, последующая обработка мазка бактерий спиртом вызывает суживание пор в пептидогликане и тем самым задерживает краситель в клеточной стенке. Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10 % массы клеточной стенки); они обесцвечиваются спиртом и при обработке фуксином или сафранином приобретают красный цвет.
В состав клеточной стенки грамотрица-тельных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана (рис. 2.4 и 2.6). Наружная мембрана при электронной микроскопии ультратонких срезов бактерий имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплаз-матической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов.
Наружная мембрана является мозаичной структурой, представленной липополисахари-дами, фосфолипидами и белками. Внутренний слой ее представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Таким образом, наружная мембрана асимметрична. ЛПС наружной мембраны состоит из трех фрагментов:
липида А - консервативной структуры, практически одинаковой у грамотрицатель-ных бактерий;
ядра, или стержневой, коровой части (лат. core - ядро), относительно консервативной олигосахаридной структуры;
высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями.
ЛПС «заякорен» в наружной мембране ли-пидом А, обуславливающим токсичность Л ПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кето-дезоксиоктоновая кислота (З-деокси-О-ман-но-2-октулосоновая кислота). О-специфическая цепь, отходящая от стержневой части молекулы ЛПС, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Генетические изменения могут привести к дефектам, «укорочению» ЛПС бактерий и к появлению в результате этого «шероховатых» колоний R-форм.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы с относительной массой до 700 Да.
Между наружной и цитоплазматической мембраной находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы,
нуклеазы, бета-лактамазы), а также компоненты транспортных систем.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т. е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сферо- или протопластного типа, утратившие способность к синтезу пеп-тидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия Института им. Д. Листера, где они впервые были изучены). L-формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку. L-формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембр ана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым - промежуточным). По структуре (см. рис. 2.5 и 2.6) она похожа на плазмалемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ.
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давле-
ния, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы и др.).
При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячива-ния в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплаз-матической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т. е. в процессах с высокой затратой энергии.
Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от SOS-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы - 50S и 30S. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, a 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и
зыполняют роль запасных веществ для питания и энергетических потребностей.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в красно-фиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризо-ванного неорганического полифосфата. При электронной микроскопии они имеют вид электронно-плотных гранул размером 0,1-1,0 мкм.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, пред-ставленная замкнутой в кольцо молекулой ДHK При нарушении деления в ней может сходиться 4 и более хромосом. Нуклеоид выявляется в световом микроскопе после ок-раски специфическими для ДНК методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДHK, связанной определенными участками с
цитоплазматической мембраной или мезосо-
мой, участвующими в репликации хромосомы (см. рис. 2.5 и 2.6).
Кроме нуклеоида, представленного одной
хромосомой, в бактериальной клетке имеются
вне хромосомные факторы наследственности -
плазмиды (см. разд. 5.1.2.), представляющие
собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь . Капсула -
слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бак-терий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпе-чатках из патологического материала. В чис-тых культурах бактерий капсула образуется
реже. Она выявляется при специальных методах окраски мазка по Бурри-Гинсу, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы.
Капсула состоит из полисахаридов (эк-зополисахаридов), иногда из полипептидов; например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Она препятствует фагоцитозу бактерий. Капсула антиген-на: антитела против капсулы вызывают ее увеличение (реакция набухан ия капсу лы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь - муко-идные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют глико-
каликсом. Кроме синтеза экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоп-лазматической мембраны, имеют большую длину, чем сама клетка (рис. 2.7). Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары - у грамотри-цательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
Жгутики состоят из белка - флагеллина (от. flagellum - жгутик), являющегося антигеном - так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных
веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования (рис. 2.7), более тонкие и короткие (3+10 нм х 0,3+10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они обладают антигенной активностью. Различают пили, ответственные за адгезию, т. е. за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водно-солевой обмен, и половые (F-пили), или конъюгационные, пили.
Обычно пили многочисленны - несколько сотен на клетку. Однако половых пил ей обычно бывает 1-3 на клетку: они образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F -, R -, Соl-плазмиды). Отличительной особенностью половых пилей является их взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях (рис. 2.7).
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки (рис. 2.8).
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не являет- i ся способом размножения, как у грибов.
Спорообразуюшие бактерии рода Bacillus , у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена з красный, а вегетативная клетка - в синий.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор может быть овальной, шаровидной; расположение в клетке - терминальное, т. е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены) и центральное у сибиреязвенной бациллы).
Процесс спорообразованя (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора. Проспору окружают две цитоплазма-тические мембраны, между которыми формируется толстый измененный пептидогликановый слой кортекса (коры). Изнутри он соприкасается с клеточной стенкой споры, а снаружи - с внутренней оболочкой споры. Наружная оболочка споры образована вегетативной клеткой. Споры некоторых бактерий имеют дополнительный покров - экзоспориум. Таким образом формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколи-новой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойную оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: ак-
тивацию, инициацию, вырастание. При этом из одной споры образуется одна бактерия. Активация- это готовность к прорастанию. При температуре 60-80 °С спора активируется для прорастания. Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.



